- When will AI surpass Facebook and Twitter as the major sources of fake news? – May 23, 2023
- The evolution of aging – May 7, 2023
- The (new) telomere theory of aging – April 15, 2023
Kin selection and aging
As individuals age, their behavior, and role within their population naturally evolve. For instance, aging may reduce an individual’s attractiveness to potential mating partners, prompting them to shift their focus towards activities that benefit their existing offspring, or even the offspring of their offspring. Many proponents of neo-Darwinism accept the concept of kin selection, despite it being a form of group selection.
Menopause is a typical example. Obviously, damage accumulation has nothing to do with this developmental, age-dependent trait. It is widely believed that the primary cause of menopause is the gradual depletion of ovarian follicles, which are limited in number since birth and cannot be replicated. Thus, menopause is a genetically programmed hallmark of aging. The common explanation for this evolved trait is that the genes of women who provide care for their grandchildren may have a higher likelihood of contributing to the evolutionary process of future generations. Please take note that there are multiple reasons why a decline in an individual’s reproductive fitness can prove advantageous for its offspring.
According to neo-Darwinists, the reason individuals in nature do not typically reach the maximum lifespan of their species is that they often die first from various causes, such as predation and disease. Hence, there is negligible selection pressure to develop characteristics that extend lifespan beyond the age of reproduction. The so-called disposable soma theory asserts that aging has little or no impact on evolution, as it takes place after reproduction.
In my opinion, the premise of this theory is incorrect. In fact, I believe that the primary reason for death in many multicellular organisms is aging.
One argument I like is that aging is an evolutionary adaptation because it reduces pathogen transmission to offspring. Older individuals are exposed to pathogens for longer times, which means their likelihood to infect other individuals in their population is higher than for younger individuals. Therefore, the survival chances of offspring are improved if older individuals are removed from the population in time.
This explains why the immune system of older individuals tends to weaken. It ensures that aged individuals die more easily of infection, allowing them less time to infect others in their population. We saw a life demonstration of this evolved trait during the pandemic. The longer a pathogen survives in a host, the higher the chances are that it adapts to the immune system of the species. Reducing the average lifespan of the species reduces the risk that an evolved version of the pathogen infects the very young with untrained immune systems. Similar to how the immune system destroys infected cells in a multicellular organism to prevent the spread of infection, infected individuals are removed from their population.
This microbiological argument works in a similar fashion at the macrobiological level. It is a well-known fact that predators always go after the weakest individuals in a group of prey. And who are the weakest? Right, the youngest and the oldest. This is the reason aging reduces physical fitness. It creates a different hierarchy of the weak than in an immortal species. In an aging species, the very young are no longer at the bottom of the weakness hierarchy, especially if you consider the extra protection they receive from their parents and the entire group.
Yes, aging improves the survival chances of grandchildren if grandma is sacrificed to the saber-tooth tiger. This is also true if grandma would still have had many good years ahead. Thus, even though grandma didn’t die of old age, she essentially died of aging when she served as breakfast for the saber-tooth tiger. The biological function of grandma’s sarcopenia is to slow her down when her offspring tries to escape a predator.
Another argument you probably heard before is that aging ensures that scarce resources only allow a limited population size. Looking at it from an evolutionary standpoint, it makes sense to feed babies instead of grandmas because the novel gene combinations among new members of the population provide fresh material for natural selection to act upon. Hence, aging is an evolved trait that reduces individual fitness to improve the survival chances of young kin. Evolution is nothing else than the replacement of the old with the new.
The main logical flaw of the disposable soma theory and its derivatives like antagonistic pleiotropy is simply that the soma is not really disposable if you assume that fitness is all about replication. The energy (food) and time required to build a new soma (alias offspring) are obviously much more costly than keeping the existing soma alive. The risk that offspring never reaches the age of reproduction is very high. Thus, to maximize the reproduction of an individual, it would be much more sensible to invest all available resources and effort in the already existing soma. Therefore, the selection pressure to evolve traits that prevent or slow aging is enormous. It can be concluded that aging serves a very different purpose in the process of evolution.
A key concept here is evolvability, which is the capacity of a population or species for adaptive evolution. Populations with a higher evolving rate evolve faster than populations with a lower evolving rate.
Evolving rate and aging rate
Species with a higher “turnover rate” of individuals have a higher evolving rate. This explains why you probably encountered a real mouse before, but never saw the slow-aging naked mole rat. Mice are everywhere. It is estimated that 20 billion of these rodents live on Earth. Yes, these vastly hyperintelligent pan dimensional beings are biologically superior by a factor of 20/8 to the species that worships the number 42 and other meanings of life beyond reproduction.
Mice are biologically fitter than slow-aging mammals because a short lifespan is an evolutionary advantage and not a handicap. Thirty mouse generations correspond to one generation of the naked mole rat if you take into account that a mouse only lives roughly one year in the wild, whereas the naked mole rat has a lifespan of 30 years. This means that in a given period, the mouse can evolve 30 times faster than the naked mole rat. Without the help of animal protectionists, long-lived Galápagos tortoises, elephants, and whales would already be extinct. In a million years, they were unable to keep up with the breakneck speed of human technological evolution.
On the other hand, these furry rodents easily infiltrated the hills built by the naked biped. As an example of adaptation, we know that city mice have shorter teeth than their cousins in rural areas. The teeth of the city mouse just have the perfect length to steal your soft cheese.

Mice adapted fast to the human threat

The naked mole rat looks like an animal from ancient times
Selection pressure and aging
The aging rate of a species is a group trait, as every species has its distinct maximum and average lifespan. Obviously, these distinct aging rates are an adaption at the group level.
The neo-Darwinist’s favorite objection against group selection is that cheating or selfish individuals have a higher chance to pass on their genes than members that exhibit altruistic behavior for the benefit of the group. In other words, the selection pressure at the individual level beats the selection pressure at the group level.
The flaw of this argument is that it assumes that individual fitness and group fitness simultaneously compete against each other. However, the time periods during which each factor operates differ significantly.
Imagine two populations of a species that don’t interbreed because of a physical barrier. The average telomere length of Population A is a bit shorter than the one of Population B which means that the individuals of Population A have a shorter average lifespan than the individuals of Population B. Thus, the reproduction rate of the individuals in Population B is higher because a longer life gives more opportunity to reproduce. However, because the turnover rate of individuals in Population A is higher, it can evolve faster, that is, the evolving rate of Population A is higher.
If one of the populations invades the territory of the other population after a significant time, say because environmental changes caused food scarcity and forced one population to migrate, a high population number in Population B faces a small population number in Population A. However, because Population A evolved faster it now has a significant advantage over Population B which can lead to the extinction of Population B. The evolving rate has turned out as more powerful than the reproduction rate and group selection determined the course of evolution.
Note that aging is only one factor that influences the evolving rate. Another important factor is the general selection pressure in an ecological niche. For instance, the selection pressure depends on parameters such as the number of competing species, the evolving rates of prey, the speed at which the physical environment changes, and many other factors. Evolution is an insanely complex, multilevel game. The important takeaway here is that the evolving rate is an important factor in evolution.
One well-known example is the invasion of Homo Neanderthalensis territories in Eurasia by Homo Sapiens. Because humans evolved faster than the Neanderthals since the two species diverged from a common ancestor around 500,000 to 700,000 years ago, the extinction of our cousins was unavoidable even though they probably were higher in number and physiologically better adapted to the region when our ancestors invaded.
In this case, it took half a million years until the selection pressure at the group level showed its power. By contrast, individual fitness is determined in a time frame of one or two generations. As mentioned above, individual fitness and group fitness operate at drastically different time scales, which the simple mathematical models of the neo-Darwinist usually neglect.
In fact, neo-Darwinists sometimes argue that the large time scales on which group selection operates is the reason selection at the individual level is more powerful. Individual selection is faster than group selection, so to speak. However, there is plenty of empirical evidence that this argument is false. I outlined one example above. The physical separation of populations also separates individual and group selection for large periods of time. Group selection strikes when groups with different evolving rates clash after hundreds of thousands of years.
Individual immortality and species mortality
What do you believe is the species that is more likely to face extinction in the near future? The fast-aging mouse or the slow-aging naked mole rat?
Only a few species are left that have not evolved the ability to age. Isn’t it interesting that only relatively primitive species, such as jellyfish or flatworms, don’t age, whereas higher-evolved species do? This is an obvious contradiction to neo-Darwinism and the disposable soma theory of aging because if aging causes little selection pressure, non-aging species should only appear much later in evolution (if at all). And what happens to all the accumulated damage created by the wear and tear of metabolism in lobsters? How did these ancient creatures evolve these remarkable damage-repair abilities hundreds of millions of years ago? Neither neo-Darwinism nor the damage theory of aging can explain the most trivial data we have about the evolution of aging.
I am not a big fan of the little computer games that neo-Darwinists like to play to simulate evolution. There are just too many variables, and the timespans involved are hundreds of thousands of years. The complexity is mind-boggling; running a computer simulation of the Bing Bang is a trivial child’s game compared to that. However, if you include aging in these models, you get exactly what I described above. Aging accelerates evolution, and immortal species have a significant fitness disadvantage that eventually leads to their extinction.
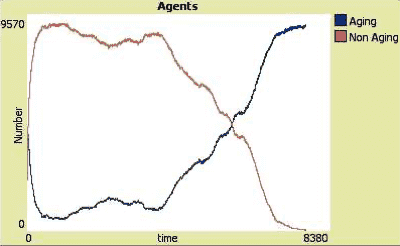
In computer simulations, immortal species get extinct if they compete against an aging species
In the curves above, you see that in this computer simulation, the gene pools of the two species have different evolving rates. The evolving rate of the non-aging species is lower, which eventually leads to its extinction when competing against an aging species with a higher evolving rate.
Neo-Darwinists focus too much on replication rates and neglect evolving rates. Obviously, a trade-off exists between those two parameters. A long lifespan with its corresponding high replication rate is at first helpful for the fitness of the species but becomes harmful if non-aging individuals keep adding their gene combinations to the gene pool for too long because their genes are no longer the optimal solution for a changed environment.
If competing species or prey evolve faster, it is only a matter of time before the once high replication rate of the individuals of a species decreases and natural selection deselects an entire species. The gene pool of the species then stops “replicating.” Yes, not only the individuals (the somas) can be seen as replicators, the gene pools of different species replicate and compete as well. The higher the turnover rate of individuals, the faster the old can be replaced with the new, the faster innovations will occur in the gene pool which corresponds to a higher evolving rate giving a population or a species a competitive advantage in the long run.
Geroscience and group selection
It appears to me that many geroscientists missed the latest development in evolution theory. Aging as an evolutionary useful trait contradicts their grad school knowledge about group selection and Darwinism. Papers that openly discuss the idea that aging is genetically programmed are rarely published in high-impact journals. The aged scientists who are in charge of these journals and who are not experts in evolution theory still consider the mere mentioning of group selection to be taboo.
Thus, many geoscientists are struggling to explain how an organism’s biological age can be reprogrammed through just a few transcription factors. It is evident that anything that can be re-programmed must have been programmed in the first place. And if aging is programmed we no longer need the disposable soma theory or antagonistic pleiotropy, excuse theories that scientists only made up to justify that all cellular and systemic damage is not simply fixed by the powerful repair mechanisms all lifeforms possess.
Conclusion
After studying the history of science for some time, I came to the conclusion that whenever mathematicians take over a field in the natural sciences, nothing good will come out. The most prominent example in physics is string theory. Lots of “beautiful” math, no empirical evidence. Neo-Darwinism is the string theory of biology. Their simple mathematical models fail to explain the most evident empirical data in evolution, aging just being one of them. I recommend reading Josh Mitteldorf’s excellent overview of the most prominent shortcomings of neo-Darwinism.
If group selection is an important factor in evolution or not is only of academic interest. In contrast, the outcome of the work of geroscientists is of fundamental importance for all of us. This is why it really bothers me that geoscientists base their theories on this hopelessly oversimplified and outdated theory of evolution that neo-Darwinism proposes. It is possible that a cure for aging could have been available for decades if geroscientists had acknowledged that aging is a programmed developmental process.











Aging is definiely programmed into us imo. How the proteome has large changes according to a study at approximately 34, 60 and 79 years of age is interesting also. It seems to kind of align with when a lot of people claim they started really feeling the aging.
It is true that the proteome programmatically changes with age. However, I think we have to distinguish between the occurrences of the typical signs of aging and the underlying biological process. Aging starts 4.5-10.5 days after the sperm and the egg cell unite and from then on the aging process decelerates until it comes to a complete stop.
This is from Steve Horvath:
Take hair graying as an example. Recent research has revealed that the discoloration of hair is not caused by the natural deterioration of melanocytes, which are responsible for pigment-producing hair. The change in behavior of melanocyte stem cells is the actual cause, and it could be connected to the programmed elimination of stem cells. The process of hair graying actually begins long before the first strand even appears, and is determined by a genetic program. When your hair turns completely white, this programmed aging process can be considered complete.
It has been evident that the theory of aging based on damage cannot fully account for hair graying, as there are significant variations among individuals in terms of when this process begins. This article explains why scientists have long overlooked the most apparent empirical data.
I guess most people would think as aging kicking on in their mid-30s if they are in decent shape because that is the point most physically decline even if they continue to exercise. I see your point on when it really starts though.
I do wonder what is limiting lifespan (in mice/rat experiments) though beyond the signaling of proteins etc. With enough age does lipofuscin and other things that are extra cellular cause enough problems as to assure death even if most factors in the blood are signaling for more youthful repair. The plateau in DNAm age is interesting also.
Are 110+ year old people mostly just dying because of stem cell exhaustion (not sure if this is the right term) that results in their hearts giving out basically or could something else be adjusted/treated so that the heart continued to replace the cells it needed to function.
Many individuals previously held the belief that the Earth was flat and that aging was a result of the accumulation of damage simply because those were the observations made by their senses. It required significant effort from scientists to persuade them otherwise.
Multicellular organisms die of old age when a vital organ fails because damage is no longer repaired. There are several reasons why systemic damage may not be repaired, with programmed stem cell elimination being one of them.
I do agree the old way of thinking about aging held up progress. If not for that we likely would have approved treatments already fine tuned that result in much better healthspans and we would be a lot closer to figuring out a way to keep people from ever reaching the current state of old age in the first place. Not sure if I will see it in my lifetime but I think once we figure it out there will be no age limit. We will go from our limit of around 120 to death from old age being eliminated. I dont buy this stuff about the age limit being first increased to 150 and 200 etc.
Do you recall when experts cautioned that it would take a minimum of 10 years to create a new vaccine at the onset of the pandemic? It’s common for experienced individuals to rely solely on their past experiences, disregarding any new factors. Given the global impact of the virus and the significant amount of funding allocated towards finding a vaccine, it was evident that the prediction of a 10-year timeline was entirely baseless.
It is possible that a breakthrough in finding the cure for aging could occur suddenly, similar to how advancements in AI have emerged overnight. Based on the substantial funding that has been dedicated to this research over the past few years, I am optimistic that a cure will be developed within the next 5-10 years.
I hope you are right. Money and the will to do it seem to be in place now and with AI assisting you may end being right. Until then I am just doing some basics like taking a large amount of liposomal Fisetin every 6 months or so and doing some plasma donations to remove some bad stuff every year. I also take calcium akg and niacinamide. I am 54 now but since I started those things my knee inflammation went away and my blood readings and blood pressure improved.
https://www.biorxiv.org/content/10.1101/2023.08.06.552148v1.full
It appears Katcher’s group has revealed its mostly the exosomes in the young pig blood that is reversing the epigenetic age. There was also another study recently that said when they removed the extracellular vesicles from the blood they didn’t get the beneficial effects they did when they remained in the transferred young blood. It would lead you to think this should work for people also and give a big healthspan effect with some lifespan extension.
Thanks for the link. For some reason, this paper did not appear in my channels. I am pretty disappointed. It seems Katcher’s miraculous E5 is nothing else than the exomes. I doubt they will get a patent for a procedure that does nothing but extract the exomes.
Do you have a link to the second paper you mentioned?
https://www.nature.com/articles/s41598-023-39370-5
Here is that link to that study. I also remember a paper that just focused on the muscles and it claimed young EVs increased Klotho and some other things there.
Hi Mike, thank you for sharing the article. I found it to be quite interesting. However, I didn’t notice any mention of the removal of extracellular vesicles from the blood.
It’s unfortunate that neither of the papers discussed the objections of leading scientists in the field, such as Conboys. According to their research, dilution of pro-aging blood factors leads to rejuvenation. To confirm this, instead of injecting a saline solution in the control group, the researchers should have checked whether plasmapheresis leads to the same rejuvenation effects.
On a separate note, I found the in-vitro experiment with young extracellular vesicles in the Nature paper to be quite intriguing. I wonder how the Conboys would explain this effect.
I heard a while back the Conboys were doing a study now where they will also record the epigenetic age after treatment. I doubt I can find it again but in China they claimed they got about a 20% reduction in methylation age on some organs with plasma dilution on mice. I still think the exosomes probably have a bigger effect but diluting first might give a little more effect possibly.
It is quite evident that exosomes play a significant role in programming the cellular age. However, it is uncertain whether they contain anti-aging factors.
In order to maintain homeostasis, multi-cellular organisms attempt to keep the concentration of each component at a specific level. Therefore, adding exosomes from young animals might reduce the number of exosomes with pro-aging factors which explains the rejuvenation effects measured in both studies.
It is highly likely that exosomes with pro-aging factors exist due to the fact that senescent cells also release exosomes.